I received an email asking some common questions about polystrate fossils. To my dismay, I realized I had never actually answered these questions in writing anywhere.
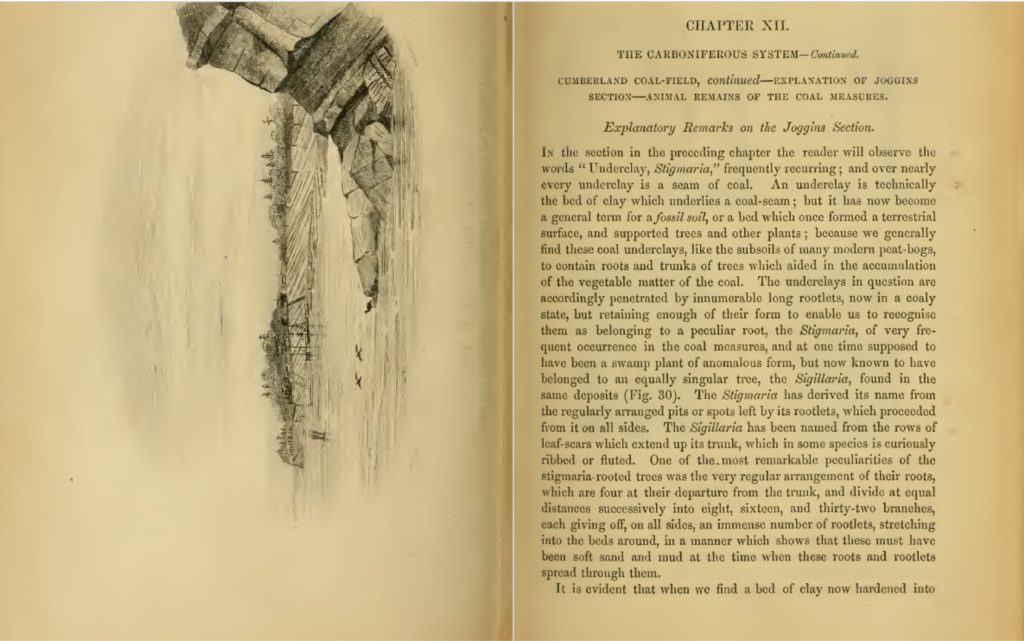
Polystrate fossils are fossils which are buried in the sedimentary record cutting through more than one sedimentary layer. The rock layers are called ‘strata’ and the fossil cuts through more than one, hence the name ‘poly’ for many and ‘strate’ for the strata the fossil cuts through. Polystrate fossils are found literally all over the world.
This fossil plant (a type of plant called a Lycopod) buried vertically cutting through layers of rock at Joggins, Nova Scotia is a classic example of a polystrate fossil.
Polystrate fossils create an impossible situation for advocates of deep time. Clearly the rock layers have not been laid down over millions of years as the top of the trunk would have rotted away long before the layers were deposited. Yet the father of deep time, Sir Charles Lyell, still advocated that these plants were buried where they grew (‘in situ’) in order to bolster his ‘new history’ of deep time he invented to replace the Biblical story of a global flood. A gentleman by the name of Paul wrote in some questions regarding polystrate fossils:
Dear Mr. Juby,
I am writing with a question about polystrate lycopod fossils. I was tipped off that I should ask you about these because I saw an image on creation.com of one such specimen which had been attributed to you. I have searched through a bit of creationist literature and have thus far not been able to find any decent scientific sources to refute the skeptic’s claim that these polystrate tree fossils must have been buried in situ because they have intact root structures that grew into the surrounding strata. I have seen a couple of creation articles claim otherwise, but I didn’t find any suitable references to back up this claim. On the contrary, I was provided by a skeptic with a reference to a very old book from the 19th century that purports to describe them thus: “One of the most remarkable peculiarities of the stigmaria-rooted trees was the very regular arrangement of their roots, which … divide at equal distances … giving off, on all sides, an immense number of rootlets, stretching into the beds around, in a manner which shows that these must have been soft sand and mud at the time when these roots and rootlets spread through them … the roots found in them were not drifted, but grew in their present positions;”
Do your own observations contradict this description? Is this description even referring to the same thing as what creationists are referring to when they talk about polystrate trees? Wading through this mess is proving difficult, so I would be glad to receive any clarification or help you might be able to provide.
Acadian Geology, John William Dawson, Third Edition, MacMillan and Co., London, 1878
Excellent questions that I hear fairly regularly. It is true that there is often intact roots found attached to these fossil stumps. I have photos of dozens of such cases. Lyell’s argument (as well as Dawson and the modern day deep time advocates) is that the intact roots are evidence that they were buried where they grew. The fact that in general, the stumps with intact roots tend to be found on the same clay bedding plane further bolstered the claim that the bedding plane was the original forest floor which was also buried in place along with the trunks.
In the photo below you can see a chunk of one of the roots (commonly called stigmaria) and you can see the pock marks around the outside, each of which radiates a rootlet. The rootlets are visible in the counter slab as black marks as the roots, rootlets and plants have all now turned into coal.
It should be noted that these plants are still around today as the Lepidodendron. Today those plants grow to maybe 16 inches tall, yet in the fossil record they are found up to 120 feet tall. The roots and the trunks are actually hollow, although the trunks had a pithy core which was displaced and infilled with sediments when the trunk was buried.
The stumps are sometimes found with the rootlets and/or roots stripped from the trunks. Clear signs that the trunks were ripped up by water, transported a distance and buried, refuting the idea of ‘in situ’ burial. But let us remember that polystrate fossils are a global phenomenon that includes far more than just the lowly Lycopods. For example, polystrate fossil coprolite (fossil poo!) has been reported at the public fossil dig sites in the Green River formation of Wyoming. Solid wood polystrate fossil tree trunks have been found in multiple places like Yellowstone National Park, or Axel Heiberg Island in the Canadian arctic. Interestingly, the ‘fossil’ stumps of Axel Heiberg Island aren’t fossilized – you can cut them with a hand saw and burn them. I have samples of this wood in my museum collection and these samples were included in the Carbon 14 testing conducted by Thomas and Nelson(1) and low and behold, there was lots of C14 found in them. But I digress. In the entire, exhaustive report by the Geological Survey Of Canada(2) on the alleged ‘in situ’ fossil forests of Axel Heiberg, you are hard pressed to find any stumps with intact roots! You will find one stump with roots extending a whopping 3 feet to a root end which has clearly been broken off. Yet these stumps are referred to consistently as buried ‘in situ.’ This is what is typically found in these alleged ‘in situ fossil forests’ claimed by advocates of deep time, so clearly their definition of ‘in situ’ and ‘intact roots and root systems’ are quite a bit different from mine!
Furthermore, the claim that the trunks of Joggins are all rooted in clay horizons (as implied by Dawson in Acadian Geology and reiterated by many deep time advocates) is demonstrably false. At the Joggins fossil cliffs the roots will be found in just about any form of deposit: clays, sandstones, shales, even coal seams. The polystrate Lycopods at the Wartburg, Tennessee coal mine were ‘rooted’ in the same horizon which was a coal seam. The polystrate trunks found in the Grande Cache coal mine (which we’ll look at closely in just a moment) were also rooted in the same horizon, a coal seam.
But let’s give the skeptics the benefit of the doubt: let’s assume that all of the stumps have intact, complex root systems in the same horizons. The roots of multiple trunks sharing the same horizon is actually not good evidence for in situ burial. Especially when one takes a closer look at those roots and their alleged ‘forest floor.’
Clearly visible roots apparently ‘floating’ above what would have been the soil line. The ‘forest floor’ horizon is marked in green.
Taking a look at the dramatic Lycopod stump, I’ve highlighted the very long root attached to the stump, complete with rootlets. Notice that the root descends from the stump, then ascends, bending upwards as if floating, and even coming up above what is claimed to be the forest floor horizon! This particular specimen has all the qualifications to meet the definition of ‘in situ’ put forward by the deep time advocates: It has intact roots and rootlets, it shares the same horizon with other stumps and roots, and it was rooted in what is interpreted as a clay horizon. Yet it is unlikely that the roots would have grown in this way – the roots grow down, not up. However, if this stump had been dislodged and transported by water, the roots would have been buoyed up in the sediments and water, thus we have an excellent explanation for this bizarre phenomenon. But – it gets better. The roots of that stump are actually entangled with the roots of a second stump which is underneath the first. The second stump is buried upside-down!
I have marked the prominent roots from the upright and inverted stumps. The inverted stump has actually fallen out of the cliff, leaving behind its impression and its root with intact rootlets. The upper stump has a second root coming down (in behind the word “roots”) which was apparently entangled with the roots of the inverted stump.
This second, inverted stump, also has all of the qualifications for the ‘in situ’ definition: intact roots and rootlets, sharing the same horizon with other stumps and roots, and it was rooted in what is interpreted as a clay horizon. But last I checked, plants don’t grown very well upside-down.
While not common at Joggins, I’d probably estimate about 1 in 100 of the polystrate Lycopods are buried upside-down. I am not the first, nor the only one to report these inverted stumps. Geologist John Mackay(3) has found some as well as Harold Coffin who documented some and provided photos of in his excellent book “Origin by Design.”(4)
When I visited the Grande Cache coal mine in 2010 to investigate the copious numbers of fossil dinosaur tracks found there, the young geologist and I made an unexpected discovery. The stratigraphic horizons are tilted up now from tectonic upheaval, almost to the vertical. We kept seeing these strange, star-shaped depressions in the layers.
When we came across one at ground level, we were shocked to find that these were all coalified stumps buried upside-down with their roots coming up into the coal seam (which had been mined away) and the trunk itself going downwards, cutting polystrate through the layers containing the trails of dinosaur tracks.
Telephoto image of one of the inverted stumps. Notice the fossil wave ripples in the sediments.Multiple, inverted, polystrate stumps had their roots in the same horizon, now a coal seam which had been excavated for the coal.One of the inverted stumps at ground level. While I couldn’t identify the plant, it was hollow like the Lycopods.
These stumps (the numbers of which I didn’t count – but as you can see were more than just a few) again fit the stipulations of the deep time advocates for interpreting these stumps as buried in situ: there were intact roots (though they have been removed in the mining operations) attached to the stumps and shared the same horizon with many other stumps. The only problem being, the trunks are all buried upside-down. Clearly these trunks weren’t buried where they grew. It’s also pretty clear that the multiple levels of dinosaur trails were not seasonal migrations as the stumps would have been sticking up from the sediments, upside down, for at least years while the sediments deposited every year for the dinosaurs to walk on. But I digress.
There is another significant argument to be made against deep time and the idea that these multiple horizons of fossil trunks were years of forest growth and burial. Take a look at this lycopod root:
A Lycopod root protruding from the cliff face.
This particular root is interesting for a few reasons. You can see the darker, differentially coloured sediments surrounding the root, as if the rootlets had clumped some soil surrounding the root when it was ripped up by water, transported and buried here. But this root, like all the others here, also tells a story of incredible pressures. The Lycopod roots were hollow and originally round. Notice the root has been squished vertically. The roots were very strong, hollow tubes and almost invariably have been crushed to failure. Although it’s difficult to see in the photo, the roots have not only been flattened vertically, they are actually split from the pressure on the top and bottom.
The sedimentary layers of Joggins total almost 20,000 vertical feet. You can walk miles of beach and because the layers have been tectonically tilted to the south (and then planed off flat across the top – see my CRSQ paper on these Planation surfaces(5)), you can examine thousands of vertical feet of sediments from the comfort of a stroll down the beach.
You’ll find roots throughout the thousands of feet which have all been compressed vertically. You’ll also find fossil logs which have been compressed to incredible levels, such as this one:
One of thousands of compressed and petrified fossil trees. Interestingly, the hollow plants tend to be buried polystrate (unless they were flattened) while the solid wood trees tend to be buried prostrate. Notice the log has been flattened vertically from compression at about 4:1 thickness ratio.
The pressures required to do this are astonishing. We can draw several, simple conclusions: 1) the trees and roots were still green when they were compressed. 2) the trees and roots had not rotted (they are preserved in pristine condition). 3) The trees had not yet petrified when they were compressed because simply put, you can’t compress a rock! 4) The Lycopod trunks and roots all show bending and compression which had to have happened while they were not yet turned to coal – they bent, instead of fracturing and disintegrating as hard and fragile coal would do. 5) The sediments the trees and Lycopods were buried in had not yet lithified (turned to solid rock) as the sediments themselves show compression and deformation along with the plants.
Everyone agrees that the compression was caused by the weight of the enormous overburden of sediments piled on top of the plants. In experiments conducted separately by myself and Vance Nelson (Creation Truth Ministries, Alberta, Canada) we were unable to deliver sufficient hydraulic pressures to mimic the compression we see in the fossil record. In my own experiment, I had pressures equivalent to hundreds of feet of overburden. So let’s play it safe with an unrealistically conservative number and say it requires a mere 300 feet of sediments to crush the Lycopodium tubular roots and the tree trunks. Starting at the bottom of the Joggins formation, we find crushed fossil logs. So a minimum of 300 feet of sediments need to be rapidly piled on top of the log before it rots, while it is still green and compressible, before it turns to stone, and before the sediments turn to stone. But in that 300 feet of overburden you will find many more crushed, petrified and coalified tree trunks and Lycopod roots – all of which also had to have been rapidly buried under a minimum of hundreds of feet of soft sediments before the plants rotted, while they were still green, before the plants turned to coal or stone, and before the sediments turned to solid rock.
But in that 300 feet of overburden sediments required to do this, you will find more crushed logs and Lycopod roots. Ad infinitum. You can put the entire ~20,000 vertical feet of sedimentary layers of the Joggins Formation into one, rapid burial event that accumulated so fast that the plants buried within were still green, had not rotted, had not turned to stone or coal, and the sediments had not yet had a chance to turn to solid rock.
Oh, and did I mention this is consistent with every formation containing polystrate fossils you find around the world?
What kind of event would rapidly bury millions of plants in tens of thousands of feet of sediments over a provincially sized area? I can assure you of one thing: whatever event that would be, we would call it a catastrophe, and on a global scale – not long and slow geological processes. Interestingly, that planation surface at the top of the cliffs is also a pertinent part of the big picture: the layers were eroded by huge volumes of extremely fast moving water. Everything about the Joggins formation in particular (but also attributable to every other polystrate fossil formation you’ll find) speaks of a huge, watery catastrophe of global proportions. I would contend it was the flood of Noah’s day. Yes, there is evidence for Noah’s flood. It’s all around you, and it’s profound and overwhelming.
November 19th in the thriving metropolis of Sioux Lookout, Ontario!
Program 1 in the Robotics: Learn by building series:
Introduction to Electricity and Electronics
I’ll be hosting the first program in the “Robotics: Learn by building” series of in-class courses. Learn electricity and electronics with no prior knowledge or skills needed!
Youth program November 19th, 9am-4pm
Suggested age 9-16: In this intensive, full day program you will learn the basics of electricity and electronics by building a number of electronic circuits using professional electronic engineering tools. No prior knowledge (except very basic math) or skills needed. Parent/guardian participation is encouraged (and they get in for free!), so drag a parent or guardian along with you! You will make your own electronic components, and use microchips to flash lights, make sound effects and control a DC motor and a servo motor, all with a focus on electronics for robotics. You can purchase all the needed parts in a kit to use in the program, or bring your own electronic parts if you have them.
There will be a treasure hunt with prizes during the breaks throughout the day.
Make sure to pack a lunch and some snacks – you’ll be using your brain extensively which consumes a lot of energy.
The Robotics: Learn by building series of programs walk you from zero knowledge in robotics to actually designing and building your own robots from scratch. This present program is the first in the series of in-person programs with extensive hands-on learning. While the program is geared for the 9-13 year old range, Ian has had 8 year olds to 40-somethings take the program and enjoy it.
The accompanying electronics kit is optional, as you may have the parts already. You are welcome to bring your own parts, just make sure you bring all of the parts in the parts list(click the big blue button to download the list)
Parent/Guardian participation with their child during the kids program is encouraged, but not necessary. Please provide an email address or contact info or download the information form and fill out one for each child.
Student seating is limited to 10 seats per class, so sign up early! A $40 deposit is required to reserve your seat. The programs will be held in the meeting room at the Best Western, Sioux Lookout
You can pay right here online using the convenient paypal link down below, or sign up by contacting Ian through email: ianjuby@ianjuby.org or telephoning toll-free at 1-877-532-9160 and hit “1” when you get the menu.
An excellent question. I was getting there in my presentation, but alas, I ran out of time. TDG was very gracious and even acknowledged that I did only have so much time to present my case. This is a question Creationists sometimes get razzed for.
A couple of comments are pertinent first.
Creation and Evolution exhaust all other possibilities:
As I stated in my opening introduction, the theories of Creation and Evolution really do exhaust all other possibilities. Even the idea that perhaps aliens transplanted life here on earth, or guided the evolutionary process here does not answer the question “Where did life come from?” It merely brings up the question “Where did the aliens come from?”
Either we were created by a being who is not only outside of our natural laws, but created those natural laws – OR – we evolved through extranatural processes which defied known natural laws.
There are no other plausible arguments for the origin of life. Therefore, an argument against evolution is, in a general sense, an argument for creation.
We need to look at the evidences to see which theory has more credence. You cannot “prove” it either way, really. TDG’s question is a pertinent one.
Proof of a Creator:
Take a look at this chair.
Let’s ask some questions:
-Did this chair have a designer?
-Did this chair have a creator?
I pointed out the irreducible complexity of both living cells and the bacterial flagellum. Those who have stumbled onto this webpage and haven’t got a clue what I’m talking about, are welcome to read the flagellum argument here.
Creationists frequently get ridiculed for not having enough blind faith. The “argument of incredulity” is definitely a valid argument.
Judge for yourself: Is it more scientific to conclude that this chair was formed by random, natural processes without any guidance, or that it was deliberately fashioned?
Let’s test the hypothetical answers with the Hogwash-O-Meter®:
“This chair was formed by natural processes without any intelligent influence.”
Oh, MAN! Another
Hogwash-O-Meter bites the dust.
One look at this chair, and one immediately recognizes the evidence of intelligent design! It is a perfectly logical, scientific conclusion to assume it had a creator, though we’ve never seen that person, nor did we see them make it.
This is not circular arguing. Saying “it exists, therefore it must have been created” is circular reasoning, just as saying “It’s here, therefore it must have evolved” is too.
We are looking at an object and deducing that it was fashioned with design and intelligence, and not randomness. In fact, we are using intelligence to deduce intelligence! It has to be the highest of irony (and hypocrisy?) that the strongest antagonists towards the Intelligent Design movement are also some of the biggest proponents of the SETI project: Searching outer space for signs of intelligence in radio waves potentially from alien lifeforms who must have evolved in our huge universe! They deny that intelligent design points to an intelligent creator (of life), then go looking for an intelligent creator (of radio transmission equipment) by looking for intelligent design in radio waves!
It is an infinitely simpler task to form this chair by natural, random processes than it is to form even the simplest of lifeforms – so if it is so impossible for this chair to form by natural processes, why on earth would a perfectly intelligent person conclude that life was formed by natural processes?
Not knowing the creator of the chair does not mean it did not have one. Not seeing him does not mean he does not exist. Perhaps that creator wants you to chase him down? Perhaps that creator wants to give people the freedom to ignore him and reject him… or chase after him? The creator is not going to force himself on you, but the evidence of him is all around you.
The argument of Homologies:
Incidentally, a common argument in favour of evolution is homology. Basically, when one looks at different animals, you can see similarities in the skeletal structure between them. This is considered evidence that they evolved from a common ancestor. I agree that this could be interpreted that way, however this is (at best) a weak argument. For example, there are plenty of chairs around, and they frequently look the same! This is just as much an evidence of common design, (i.e., it works well, use it in other designs!) just as much as it could be argued they evolved from the same ancient chair.
Homologies are left in the dust however, when one brings the duckbilled platypus into the picture. These arguments of homologies are pertinent to the “Suspected references” response.
Radiohalos: Evidence of an instantaneous formation of earth
The next part of my opening talk would have included the evidence in geology for a young earth, and an instantly created one. Rather than re-invent the wheel, I will merely point readers to my already published webpage on the radio halos in granites. This is irrefutable evidence of an instantaneous formation of earth’s granite crust, and presumably, earth itself.
In brief, the granite crust of the earth has no natural origin. Anyone who would beg to differ is quite welcome to answer this challenge by merely producing synthetic granite in the lab. It IS physically impossible. Furthermore, the granite we do find shows evidence of instantaneous formation.
This happens to be a subject I am very knowledgeable in and about which I am quite willing to field questions and detractions.
The Nowosiolka skull considered as a proof of the existence in recent history of forms belonging to H. primigenius.
Memoir of M. Kaimierz Stoly who presented by M. H. Hoyer on the 7th of January 1908.
During the last several years I published several works concerning the existence, in Poland, of a Spy-Neanderthaloid type. These works attracted the attracted the attention of the professor G. Schwalbe, seeing as the subject contradicted several of his opinions. According to Mr. Schwalbe, the Spy-Neanderthal race existed only in the ancient deluvian period, after which it went extinct, as well as the transitional forms between H. primigenius and H. sapiens did not go beyond the Paleolithic. To the contrary, I was of the opinion that the Spy-Neanderthaloid types, having a softer Neanderthal-type structure, existed not only in the ancient deluvian, but also in the later prehistoric periods and can be observed even in the historic period. Of course, there is no question of the existence beyond the Paleolithic of a pure Spy-Neanderthal type, exactly the same as the bones of Neanderthal, the Spy, and to a lesser degree, the bones of the Krapina; it consists only of the existence beyond the ancient deluvian of human bones having a structure that resembles that of the Spy-Neanderthal type, although a lot more delicate than this and even changed in some aspects thanks to the influence of the environment and growth. Mr. Schwalbe found that this way of looking at things was wrong and submitted a critique to the German Anthropological Congress of 1905, also in a memoir that appeared in the Zeitschrift f. Morphologie u. Anthropologie J. 1906. I’ve also been honored to write several letters on the subject. To solve the problems in question, I have submitted the materials previously studied to a new examination; my previous research, not being in the manner of the methods of Mr. Schwalbe did not permit comparison between my skulls and those which were considered by Misters Schwalbe, Klaatsch and Gorjanowie-Kramberger as representatives of the Spy-Neanderthal race. I started by studying use the method of Mr. Schwalbe the “scythic” cranium of Nowosiolka; it’s my opinion that it is the one amongst all the skulls to be found at the Anthropological laboratory of Varsovie that contains the characteristics the most representative of the Spy-Neanderthal race.
We have found archaeological facts concerning the skull in question in a memoir of Mr. Bydlowski “the Kourghans of Nowosiolka of the district of Lipowiec, Government of Kiev” (Swiatowit t. V. 1904); the kourghan from where the skull came is designated as number V. The skull of Nowosiolka must have belonged to a man that was thirty years old; concerning the period from which he came, we can determine this from the objects that were found with him in the tomb. Beside the skeleton, which was lying on his back, the head towards the west, was found a suit of armour complete with iron plates held together by rivets. On the other side were found two long spear points of which one had a flat side, the other was forged in a quadrangular bar. At the feet of the skeleton was a yellow clay goblet with a handle of ornate work, bordered on the inside with a black band; two parallel bands went around the inside of the goblet at the top. Beside the goblet was found a beautiful red clay amphora (antique vase) of conical shape, the bottom shaped obliquely. At the feet and the head of the dead were found two horse skeletons with the harness bridles and four bits the same as those that are used today. Concerning the harness we found only bits of leather and green and oxidized metal ornaments.
The present memoire is solely dedicated to the study of the position that the Nowosiolka skull when compared to the H. primigenius group, which includes the skulls of Neanderthal, of Spy and of Krapina, and also taking into consideration other diluvian skulls. I will not concern myself with the question of the relationship between H. primigenius and H. sapiens, nor of the problem of where the limit is to be marked between these two groups.
Tori supraorbitales. I start with this characteristic, recognized as pithicoidal and having, according to M. Schwalbe the most importance as a distinctive trait of the Spy-Neanderthal type. M. Schwalbe affirms that all developed eyebrows cannot be considered as a sufficient enough characteristic to classify a skull amoung the representatives of the Spy-Neanderthal type; The indispensable condition is that the eyebrow protrusion must follow the borders of the entire orbital (eye sockets), with the exception of a slight depression which can be found on the median plane. M. Schwalbe calls these protrusions tori supraorbitales and considers them, the same as M. Klaatsch and M. Gorjanowie-Kramberger, as the most important characteristic amoung the Spy-Neaderthal race. Concerning H. Sapiens, we may sometimes find, according to M. Schwalbe, pronounced eyebrows but which instead of following the complete orbit to the joint of the frontal and cheek bones, hardly get to the middle after which they go obliquely straight up. There exists in H. sapiens, in the lateral part of the orbital, in a trilateral plane or even of a small depression that extends from the middle of the upper edge of the orbitals to where they meet the cheek bone. The lateral plane in question is called by M. Schwalbe planum supraorbitale. For M. Klaatsch and M. Gorjanowie-Kramberger and M. Schwalbe, the protrusion of the lateral part of the eyebrows never happens in modern skulls.According to M. Gorjanowie-Kramberger, the sus-orbital cut (indentation) in the case in question is an important morphological characteristic. According to this author, Neolithic skulls and skulls belonging to recent H. sapiens types present a sus-orbital cut sometimes very deep of which the lateral edge defines the sus-orbital protuberance. Contrarily, the skulls belonging to the H. primigenius type have only a very weak protuberance and laterally a small oblong extension. In addition, the sus-orbital protuberance ordinarily gives birth to a very flat “gutter” which, laterally and obliquely directed, transverses the torus supraobrbitalis and consists of the first developmental stage of the sus-orbital line, that is to say the first degree of separation of the sus-orbital prominences; they are continuous in H. primigenius, but are completely separate in H. sapiens. However, concerning tori and incisorae supraorbitales, M. Gorjanowie-Kramberger recognizes the existence of transitional types between H. primigenius and H. sapiens and cites as an example of the gradual disappearance of the tori, the Néolithic Babsk skull.In studying the Nowosiolka skull, I was careful to take into consideration the opinions that I just cited. My research has demonstrated that 1. the skull in question has extended eyebrows that follow the complete ridge of the orbitals until they reach the cheek bones; this corresponds exactly to the characteristic structure of H. primigenius; 2. that the sus-orbital cuts are greatly developed but the gutters which go off in a obliquely lateral direction are so weak and so shallow that they do not interrupt the continuity of the tori supraorbitales. We can observe similar relationships with the Krapina skull where the gutters are very flat presenting the first degree of separation of the tori supraorbitales in arcus supraciliaris and margo supraorbitales. According to M. Gorjanowie-Kramberger this cranial structure of Krapina is evidence in favor of the existence of recent transitional forms between H. primigenius and H. sapiens. To determine more exactly the thickness of the tore orbitales of the Nowosiolka skull, I measured them and give here the results of these measurements adding also the results found by M. Gorjanowie-Kramberger concerning the tori supraorbitales of several skulls from Krapina.
(see table on page 107)
Skulls
Median edge
Point of the least thickness
Lateral edge
Neanderthal from the right
20 mm
12 mm
12 mm
Neanderthal from the left
20 mm
13 mm
13 mm
Nowosiolka from the right
16 mm
8 mm
9 mm
Nowosiolda from the left
16 mm
7 mm
10 mm
Krapina C
13.8 mm
9.5 mm
12.1 mm
Krapina D
11.9 mm
9.3 mm
11.5 mm
Krapina fragment
9 mm
Another Krapina fragment
4 mm
This table shows that the thickness of the eyebrows of the Nowosiolka skull is very close to that of the Krapina skulls. It is true that the lateral part of the eyebrows are a little less thick than their equivalents in the Krapina skulls (with the exception of one of the fragments) but, in return, the median parts are thicker. In short, the eyebrows of the Nowosiolka skull, thanks to their non-interrupted continuity to the frontal-cheek suture, also in considering their thickness can be designated as tori supraorbitales and not as arcus supraciliaris, as is also recognized by M. Schwalbe based upon a photograph that I had the honour of sending him.
Concerning the eyebrows, the Nowosiolka skull is quite different from the H. sapiens type and can be joined to the Spy-Neanderthal-Krapina group.
The interorbital diameter of the Nowosiolka skull does therefore not go beyond the dimensions of H. primigenius.
Diameter of the interal biorbital:
Spy I
116-113 mm
Krapina C
107 mm
Neanderthal
112 mm
Krapina fragment
106 mm
Nowosiolka
110 mm
Brüx skull
104 mm
The biorbital diameter of the Nowosiolka skull does therefore not go beyond the dimensions of H. primigenius.
Interorbital index:
Brüx
29.8
Krapina C
27.1
Spy I
29-28
Neanderthal
27
Nowosiolka
27.27
Krapina fragment
26.4
The interorbital index of the Nowosiolka skull does therefore not go beyond the dimensions of H. primigenius.
III. Frontal-biorbital index [frontal minima width x 100 / external biorbital diameter]
Diameter of the external biorbital:
Krapina D
126.2 mm
Spy I
123 mm
Neanderthal
126 mm
Nowosiolka
122 mm
Krapina fragment
126 mm
Krapina C
118 mm
Spy II
124 mm
Pithecanthropus
106 mm
The external biorbital diameter of the Nowosiolka skull does therefore not go beyond the dimensions of H. primigenius.
Frontal minima width:
Krapina D
100.4 mm
Tilbury
101 mm
Spy II
109 mm
Krapina C
98.5 mm
Neanderthal
107.5 mm
Podbaby
98 mm
Nowosiolka
104 mm
Cannstadt
96 mm
Spy I
104 mm
Gibraltar
95 mm
Krapina fragment
104 mm
Brüx
92 mm
Sligo
102 mm
Pithecanthropus
87 mm
The frontal minima width of the Nowosiolka skull does therefore not go beyond the dimensions of H. primigenius.
Frontal biorbital index
Neanderthal
88.8
Spy I
84.5
Spy II
87.9
Krapina C
83.4
Krapina D
87.3
Krapina fragment
82.5
Nowosiolka
85.25
Pithecanthropus
82
The frontal biorbital index of the Nowosiolka skull does therefore not go beyond the dimensions of H. primigenius.
Bregmatic angle
Cannstadt
60
Brüx
51.5 – 45.5
Brünn
54
Spy II
50.5
Nowosiolka
53.5
Krapina D
50
Krapina C
52
Spy I
46
Galley-Hill
52
Neanderthal
44
Gibraltar
51 – 50
Pithecanthropus
37.5
The brgmatic angle of the Nowosiolka skull is close to that of H. primigenius.
Frontal angle (m. g. i.):
Cannstadt
90
Spy II
70
Galley-Hill
82
Krapina C
70
Nowosiolka
78 *
Krapina D
65
Brüx
77 – 72.5
Neanderthal
62
Brünn
75
Spy I
59
Gibraltar
74 – 73
Pithecanthropus
52.5
The frontal angle of the Nowosiolka skull differs from that of H. primigenius. However, it is to be noted that closeness of this measurement with the Brüx skull which is considered by M. Schwalbe, in terms of the frontal angle measurements as being transitional between H. primigenius and H. sapiens.
Angle of frontal convexity (Angle A) (n. r. b.)
Spy I
151
Cannstadt
135
Pithecanthropus
146.25
Nowosiolka
131
Spy II
141.5
Brüx
131
Neanderthal
139
The angle (n. r. b.) of the Nowosiolka skull differs from those of H. primigenius.
Angle B (g. e. b.)
This angle is more important than the last one by which the opening is dependant, to a certain degree, of the position more of less depressed of the nasal opening.
Spy I
159
Neanderthal
150
Pithecanthropus
153.2
Nowosiolka
139
Spy
151
Krapina A (child)
134
The angle (g. e. b.) of the Nowosiolka skull differs from those of H. primigenius.
VII. Angle: lambda – glabella – inion (l. g. i.)
Nowosiolka
18
Brünn
17
Krapina C
17
Spy II
16.5
Brüx
17
Spy I
16
Galley-Hill
17
Neanderthal
15
The angle lambda – glabella – inion of the Nowosiolka skull is close to that of H. primigenius.
VIII. Convex index of the frontal arch
A: [diameter: nasal-bregma x 100 / curve: nasal-bregma]
Diameter: nasal-bregma
Neanderthal
119 mm
Brüx
115 mm
Cannstadt
117 mm
Egisheim
115 mm
Nowosiolka
116 mm
Pithecanthropus
112-96 mm
Spy II
115 mm
Spy I
108 mm
The diameter of the nasal-bragma of the Nowosiolka skull is within the range of H. primigenius. bregma
Nasal-bregma curve
Cannstadt
135 mm
Egisheim
130 mm
Brüx
135 mm
Spy II
124 mm
Neanderthal
133 mm
Pithecanthropus
120 – 100 mm
Nowosiolka
132 mm
Spy I
115 mm
The nasal-bregma curve of the Nowosiolka skull is within the range of H. primigenius.
Index A) of the convexity of the frontal arch:
Pithecanthropus
94.6
Nowosiolka
87.22
Spy I
93.9
Neanderthal
87.2
Spy II
92.7
Cannstadt
86.67
Egisheim
88.5
Brüx
85.1
The index A) of the convexity of the frontal arch of the Nowosiolka skull is within the range of H. primigenius.
Index of the convexity of the frontal arch
B): [diameter: glabella-bregma x 100 / curve: glabella-bregma]
This index is more important than the one before by which the size depends, to a certain degree, to the position more or less depressed of the nasal cavity; in the case of deep concavity at the base of the nose the median curve of the front gains in length; however this is not an equivalent of the growth of the convexity of the frontal arch.
Spy-Neanderthal
93.8 – 89.4
Nowosiolka
92.8
Krapina A (child)
90.3
The index of the convexity of the frontal arch of the Nowosiolka skull does not go beyond the range of H. primigenius. It is also to be noticed that the Nowosiolka skull contains a persistent frontal suture for most of it’s length, and that the light bulges that accompany each side contribute to produce the effect of convexity fo the frontal arch when seen on the photograph of the skull in question in it’s norma lateralis.
Glabello – cerebral index[diameter: frontal glabello x 100 / diameter frontal cerebral]Diameter of the frontal glabello:
Neanderthal
38 mm
Nowosiolka
25 mm
Gibraltar
36 mm
Pithecanthropus
24 mm
Spy I
32 mm
Galley-Hill
24 mm
Spy II
30 mm
Brüx
24 mm
Brünn
30 mm
Cannstadt
13 mm
Krapina C
28.2 mm
The frontal glabella diameter of the Nowosiolka skull is close to those of H. primigenius.
Diameter of the frontal cerebral
Cannstadt
104 mm
Pithecanthropus
80 – 95 mm
Brüx
99 mm
Neanderthal
86 mm
Nowosiolka
99 mm
Krapina C
86 mm
Brünn
96 mm
Gibraltar
82 mm
Galley-Hill
95 mm
Spy I
81 mm
Spy II
92 mm
The diameter of the frontal cerebral of the Nowosiolka skull differs from those of H. primigenius.
Glabello-cerebral index
Neanderthal
44.2
Egisheim
29
Gibraltar
43
Pithecanthropus
27.6
Spy I
41.5 – 40.9
Nowosiolka
25-26
Spy II
34.4
Galley-Hill
25.2
Krapina C
32
Brüx
24.24
Brünn
31.2
Cannstadt
18.2
The glabello-cerebral index of the Nowosoilka skull is beyond the range of H. primigenius. M. Schwalbe came to the conclusion that this index is very important and that it permits us to mark a limit between the H. primigenius and H. sapiens groups which are to date not linked by the existence of any intermediate form. However, the table below shows that the skulls of Brünn, of Egisheim and even that of Pithecanthropus consist of similar transitional forms. This fact is all the more remarkable when we consider that M. Schwalbe does not put aside the possibility of genetic development of Pithanthropus and H. primigenius. In this hypothesis, we would have to wait until Pithecanthropus develops precisely this distinctive and important trait to a degree as pronounced as H. primigenius. The previous table proves the contrary. The two facts mentioned above: the existence of intermediate forms and the reality that Pithecanthropus is of such form concerning the glabello-cerebral index, considerably diminishes the value that M. Schwalbe attributes to the index in question.
Angle of the protrusion of the part of the frontal cerebral (a. t. h.)
Spy I
155.5
Spy II
147
Pithecanthropus
154.5
Brüx
143.5
Neanderthal
151
Nowosiolka
139
The angle in question for the Nowosiolka skull differs from that of H. primigenius. The small size of this angle of the Nowosiolka skull must certainly have as a cause the median convexity of the frontal produced by the persistence of the frontal suture.
Index of the convexity of the cerebral part of the frontal[diameter of the cerebral part x 100 /curve of the cerebral part]
Curve of the cerebral part of the frontal
Cannstadt
113 mm
Neanderthal
95 mm
Brüx
105 mm
Pithecanthropus
98 – 78 mm
Nowosiolka
105 mm
Spy I
88 mm
Spy II
100 mm
The curve of the cerebral part of the frontal of the Nowosiolka skull differs from those of H. primigenius.
Index of the convexity of the cerebral part:
Spy I
97.5
Nowosiolka
92.38
Pithecanthropus
96.1
Cannstadt
92.04
Neanderthal
95.5
Spy II
92.0
Brüx
94.29
This index for the Nowosiolka skull is within the range of H. primigenius.
XII. Index of the position of the bregma
[diameter g. w. x 100 / diameter glabella-inion]
Diameter glabella-inion
Galley-Hill
204 mm
Nowosiolka
194 mm
Brünn
201 mm
Gibraltar
187 mm
Neanderthal
199 mm
Brüx
186-180 mm
Spy I
198 mm
Pithecanthropus
181 mm
Krapina D
197.5 mm
Cannstadt
174 mm
Spy II
196 mm
The glabella-inion diameter of the Nowosiolka skull is close to that of H. primigenius.
Index of the position of the bregma
Pithecanthropus
44.1
Spy II
35.2
Neanderthal
38.4
Spy I
34.8
Nowosiolka
35.56
Krapina D
31.8
The index of the position of the bregma is within the range of H. primigenius.
XIII. Fronto-parietal index
[minima width of front x 100 / maxima width of front]
Neanderthal
73.1
Brüx
71-68
Krapina A
72.5
Krapina C
66.4
Nowosiolka
72.22
Cannstadt
65.7
Tilbury
72.1
Pithecanthropus
65.4
Spy I
71.2
Gibraltar
64.2
Spy II
71.2
Egisheim
61.3
The fronto-parietal index of the Nowosiolka skull is within the range of H. primigenius.
XIV. Index of the convexity of the parietals
[diameter of the sagittal suture x 100 / curve of the sagittal suture]
Neanderthal
94.5
Nowosiolka
91.18
Krapina D
92.8
Krapina (adult)
86.3
This index for the Nowosiolka skull is within the range of H. primigenius.
Index of the parietals
[sagittal curve of the parietals x 100 / sagittal curve of the front]
Spy I
104.3
Brüx
92.6
Nowosiolka
102.26
Neanderthal
89.4-82.7
Spy II
96.7
Pithecanthropus
85.8
Egisheim
95.3
The index of the parietals are within the range of H. primigenius.
XVI. Cephalic index
[maxima width of the skull x 100 / maxima length of the skull]
Galley-Hill
205 mm
Brüx
195-190 mm
Brünn
204 mm
Gibraltar
192.5-190 mm
Nowosiolka
200 mm
Pithecanthropus
181 mm
Neanderthal
199 mm
Krapina C
178 mm
Spy II
198 mm
Cannstadt
178 mm
Krapina D
197.5 mm
The maxima length of the Nowosiolka skull is close to that of H. primigenius.
Maxima width (euryon – euryon)
Krapina D
169 mm
Tilbury
140 mm
Krapina C
149 mm
Brünn
139 mm
Gribraltar
148 mm
Brüx
135-130 mm
Neanderthal
147 mm
Pithecanthropus
133 mm
Cannstadt
146 mm
Galley-Hill
130mm
Nowosiolka
144 mm
The maxima width of the Nowosiolka skull is close to the range of H.primigenius.
Cephalic index:
Krapina D
85.5
Pithecanthropus
73.4
Krapina C
83.7
Spy I
72.2
Cannstadt
82
Nowosiolka
72
Gibraltar
77.9
Brüx
69
Spy II
77.2
Brünn
68.2
Neanderthal
73.9
Galley-Hill
63.4
The cephalic index of the Howosiolka skull is close to that of H. primigenius.
XVII. Height of the cranial arch in relationship to the glabella-inion diameter (Kalottenhöbe)
Cannstadt
105 mm
Gibraltar
85 mm
Nowosiolks
104 mm
Neanderthal
81-80.5 mm
Brünn
103 mm
Krapina D
83.5 mm
Galley-Hill
97 mm
Krapina C
82 mm
Brüx
92-85 mm
Spy I
81 mm
Spy II
87 mm
Pithecanthropus
62 mm
The height of the cranial arch in relationship with the glabella-inion diameter of the Nowosiolka skull differs from that of H. primigenius.
XVIII. Index of the height of the cranial arch in relationship to the glabella-inion (Kalottenhöbe index)
[diameter c. h. (cranial arch height) x 100 / diameter: glabella-inion]
Cannstadt
60.34 – 58.99
Gibraltar
45.4
Egisheim
55.5
Spy II
44.3
Nowosiolka
53.61
Krapina D
42.2
Brünn
51.2
Spy I
40.9
Brüx
51.1-47.6
Neanderthal
40.5
Galley-Hill
48.2
Pithecanthropus
34.2
Krapina C
46
This index of the Nowosiolka skull is outside the range of H. primigenius.
XIX. The height of the cranial arch in relationship with the glabella-lambda diameter (Lambda-Kalottenhöbe)
Cannstadt
75 mm
Neanderthal
57-54.5 mm
Egisheim
70 mm
Brüx
56 mm
Nowosiolka
66 mm
Krapina D
53 mm
Spy II
58 mm
Spy I
51 mm
The height of the cranial arch in relationship with the glabella-lambda diameter of the Nowosiolka skull differs from the that of H. primigenius.
Index of the height of the cranial arch in relationship with the glabella-lambda diameter
[diameter k. p. (height of the cranial arch) x 100 / glabella-lambda diameter]
Cannstadt
43.3-42.1
Brüx
30.2
Egisheim
37.8
Neanderthal
29.4
Nowosiolka
34.02
Krapina D
29.3
Spy II
31.3
Spy I
27.4
This index of the height of the cranial arch of the Nowosiolka skull is close to that of H. primigenius.
XXI. Lambda angle (l. g. i.)
Nowosiolka
82
Gibraltar
73 – 69
Brünn
78
Neanderthal
66.5
Krapina C
76
Krapina D
65
Galley-Hill
71
The lambda angle of the Nowosiolka skull differs from that of H. primigineius.
XXII. The angle of the opisthion (o. i. g.)
Krapina D
57
Galley-Hill
42
Neanderthal
51.3
Nowosiolka
36
Brünn
42
Gibraltar
36
The angle of the opsthion of the Nowosiolka skull differs from that of H. primigeius.
XXIII. Occipital convexity index
[diameter: lambda-opisthion x 100 / curve: lambda-inion-opisthion]
Nowosiolka
81.45
Krapina B
80.4
Other Krapina
77
The occipital convexity index is close to that of H. primigenius.
XXIV. Configuration of the zygomatico-maxilla.
According to M. Garjonowie-Kramberger, the zygomatico-maxilla suture of the Krapina C skull has a different configuration from that observed on recent H. sapiens skulls. The sub-orbital part of the cheek bone of the skull in question doesn’t extend as far towards the nose and is not as receded as modern and Neolithic European skulls. The result is that the zygomatico-maxilla suture of this skull goes vertically down and then turns towards the back. This configuration is related to the considerable development of the sub-orbital part of the maximilla which normally brings about the absence of canine dimples. However, with the Nowosiolka skull, the configuration of the zygomatico-maxilla suture is similar to that of the Krapina skull; the canine dimples are also missing. Therefore, it can be said that the configuration of the zygomatico-maxilla of the Nowosiolka skull does not differ from the same structure of H. primigenius.
XXV. The level of the zygomatic eyebrow. The position of the zygomatic eyebrow in relationship with the german horizontal constitutes according to M. Garjonowie-Kramberger an important morphological trait. In recent H. Sapiens, the eyebrows in question extend ordinarily either above the horizontal plane, either at the same level and only on rare occasions will it be found below this plane. With the Krapina C skull, on the temporal part of the zygomatic eyebrow starts at the level of the german horizontal plane but then lowers so much that the anterior upper edge of the cheekbone apophysis is found below the plane in question. M. Fairpont and Klaatsch think that a configuration like this existed on the Spy I skull. A similar structre can be found in several anthropoids: the gorilla and the chimpanzee. On the Nowosiolka skull the structure is exactly the same as that described on the Krapina C skull. Therefore, in terms of the zygomatic eyebrow on the Nowosiolka skull it is not different that that found in H. primigenius.
XXVI. Akanthion-prosthion diameter
Spy I
28 mm
Krapina C
21 mm
Krapina E
27.7 mm
Nowosiolka
18 mm
Krapina F
24.7 mm
The akantion-prosthion diameter is close to that of H. primigenius.
XXVII. Height of the inferior jaw index
[height of the jaw at the 2-me molar x 100 / height of the jaw at the symphysis]
Height of the jaw at the 2-me molar:
Krapina H
34.3-33 mm
Krapina B
25 mm
Spy I
33 mm
Krapina E
24.1 mm
Krapina I
32.2 mm
Arcy
24 mm
Krapina D
29 mm
La-Naulette
23 mm
Nowosiolka
28 mm
Mularnaud
22 mm
Krapina G
27.5 mm
Krpaina C
20 mm
Krapina F
27.4 mm
The height of the jaw at the 2-me molar is within the range of H. primigenius
The height of the jaw at the symphysis
Krapina I
42.3 mm
Krapina F
31 mm
Krapina H
40 mm
La-Naulette
31 mm
Spy I
38 mm
Krapina C
29-28.5 mm
Nowosiolka
35 mm
D’Arcy
28 mm
Krapina E
35 mm
Malarnaud
26 mm
Krapina D
33-30.5 mm
Krapina B (child)
25.3 mm
Krapina G
31.5-30.4 mm
The height of the jaw at the symphysis is within the range of H. primigenius.
According to M. Gorjanowie-Kramberger, the inferior jaw of H. primigenius is noticeable due to the considerable height of the jaw at the symphysis; numerous diluvian skulls also have some height at the symphysis that goes beyond the upper molar. M. Gorjanowie-Kramberger considers that when a similar structure is seen in recent H. sapiens, it is an atavic variation. This exists also on the Nowosiolka skull.
Index of the height of the inferior jaw:
Krapina G
92.1-88.8 mm
Malarnaud
84.6 mm
Spy I
86.8 mm
Nowosiolka
80 mm
D’Arcy
85.7 mm
La-Naulette
74.19 mm
Krapina H
85.7 mm
Krapina I
71.3 mm
Krapina B
84.8 mm
Krapina E
68.8 mm
The inferior jaw height index is within the range of H. primigenius.
XXVIII. Thickness of the main part of the symphysis of the lower jaw
Krapina I
74 mm
Spy I
68 mm
Krapina C
70.4 mm
Krapina H
66.5 mm
Nowosiolka
70 mm
D’Ochos
66 mm
This distance on the Nowosiolka skull is within range of H. primigenius.
XXIX. Anterior mandible angle
Krapina H
106
Krapina D
94
Krapina G
103.5
Krapina I
94
Krapina F
102
La-naulette
89
Krapina C
96.5
Spy I
84.5
Krapina E
95
Nowosiolka
68
The anterior mandible angle is not within the range of H. primigenius.
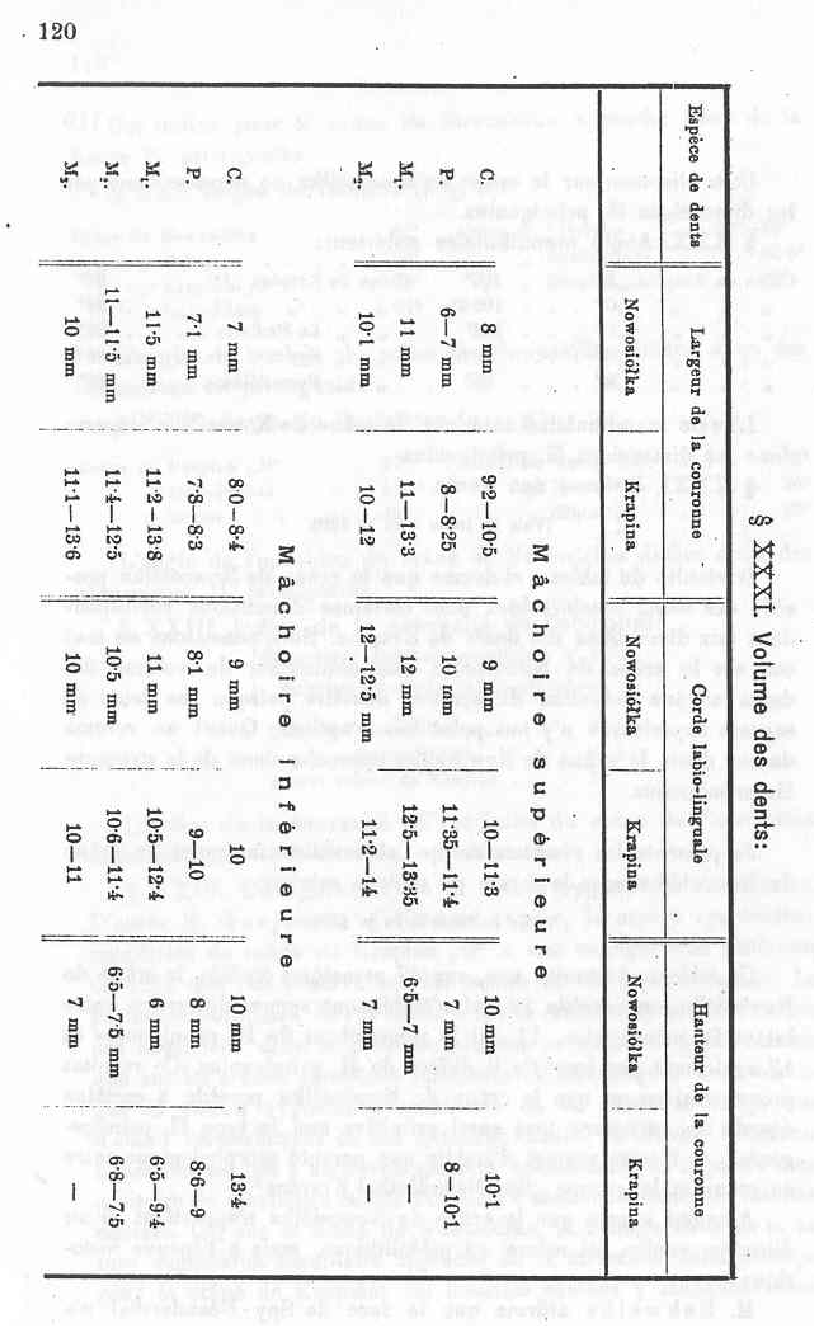
XXX. Teeth size
See table on page 120
The results from the table show that the Nowosiolka skull has some teeth of considerable size of which some correspond to the Krapina skull. We notice that in the case of the Nowosiolka skull a reduction in teeth size and a reduction in the whole dental system since the wisdom teeth have not yet appeared. Concerning the teeth size, the Nowosiolka skull approaches that of H. primigenius.
I present the results of my observations concerning the Nowosiolka skull in the following table on page 121
This table shows that, out of 47 characteristics studied, the Nowosiolka skull possesses 23 which are identical to that of H. primigenius, 11 that are close to that of H. primigenius and only 13 that differ from H. primigenius. These results prove that the Nowosiolka skull possesses in some aspects a structure as primitive as the H. primigenius type and permits us to establish a morphological link between this skull and those of Spy-Neanderthal-Krapina.
Not forgetting that the Nowosiolka skull does not belong to the ancient diluvian, nor the Paleolithic, but to recent history.
Schwalbe affirms that the Spy Neanderthal race only existed on earth until the middle of the quarterly period, period when they became extinct and replaced by H. sapiens. It is true that M. Schwalbe admits the existence of transitional forms between H. primigenius and H. sapiens but according to him, these forms to not possess the characteristic traits of H. primigenius and they probably belong to the middle diluvian because they rarely go beyond the Paleolithic.
The analysis of the Nowosiolka skull has shown that it contains numerous characteristics which are only associated with H. primigenius. Several very important, such as the tori supraorbitalis, the low level of the zygomatic eyebrow, the considerable thickness of the mandible. Also, the Nowosiolka skull according to the bregmatic angle considerably approaches that of the Brüx skull, which according to M. Schwalbe, constitutes a transitional form between H. primigenius and H. sapiens.
In conclusion, the facts cited earlier demonstrate that 1. that morphological forms belonging to H. primigenius existed not only in the Paleolithic, but also later eras as well as recent history 2. that transitional forms between H. primigenius and H. sapiens contain distinctive traits that differ from H. primigenius. IT is possible that the opinion of M. Schwalbe, that H. primigenius existed only during the ancient diluvian is true only for regions in southern Europe where the Spy-Neanderthal-Krapina type did not live beyond the Paleolithic; but this can not be applied to all of Europe. We must assume that this race was progressively pushed towards the north, where by this fact, it could survive much more longer than in the south of Europe.
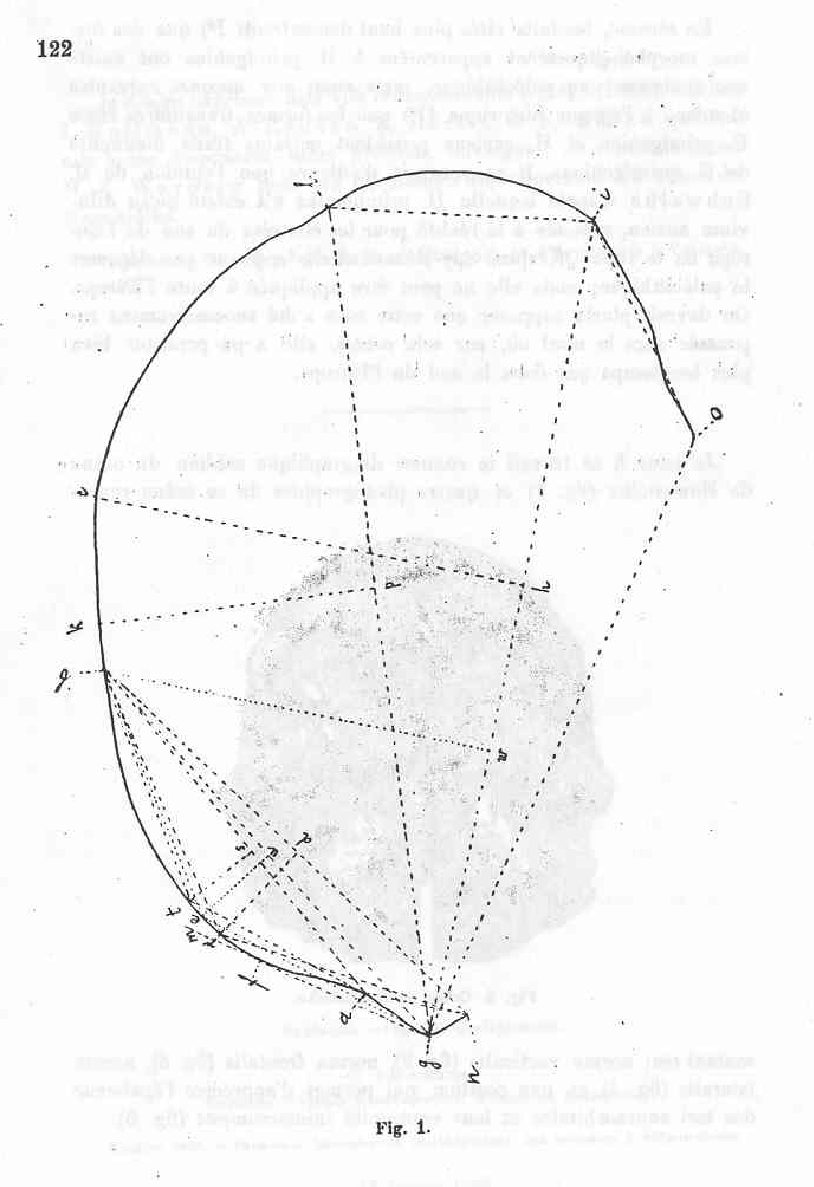
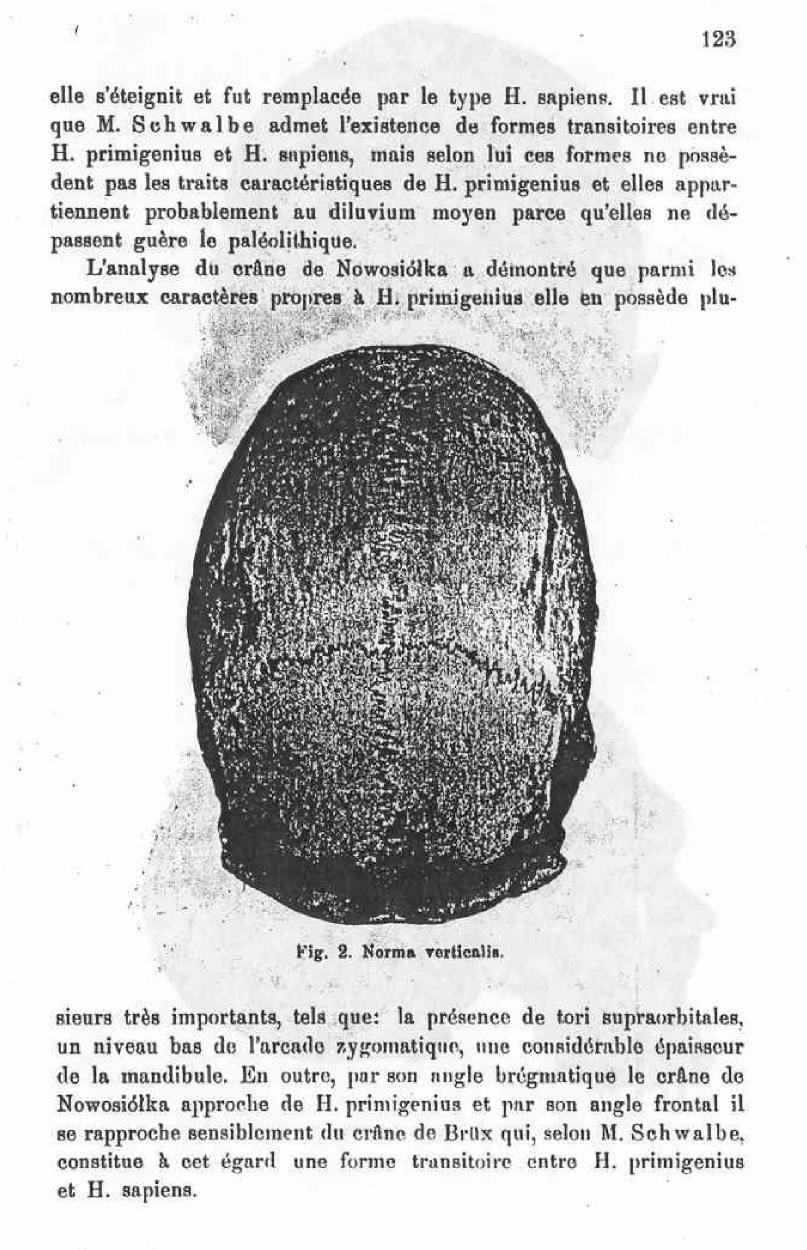
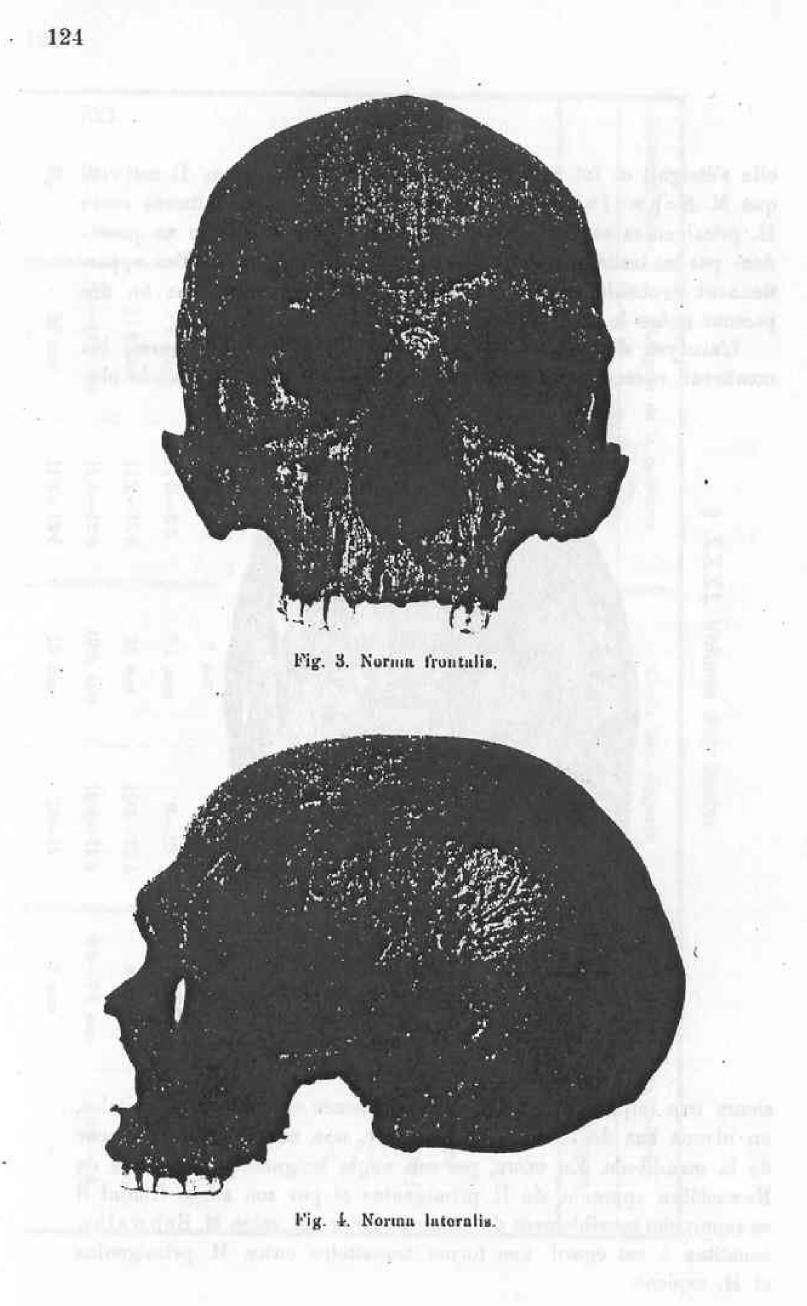
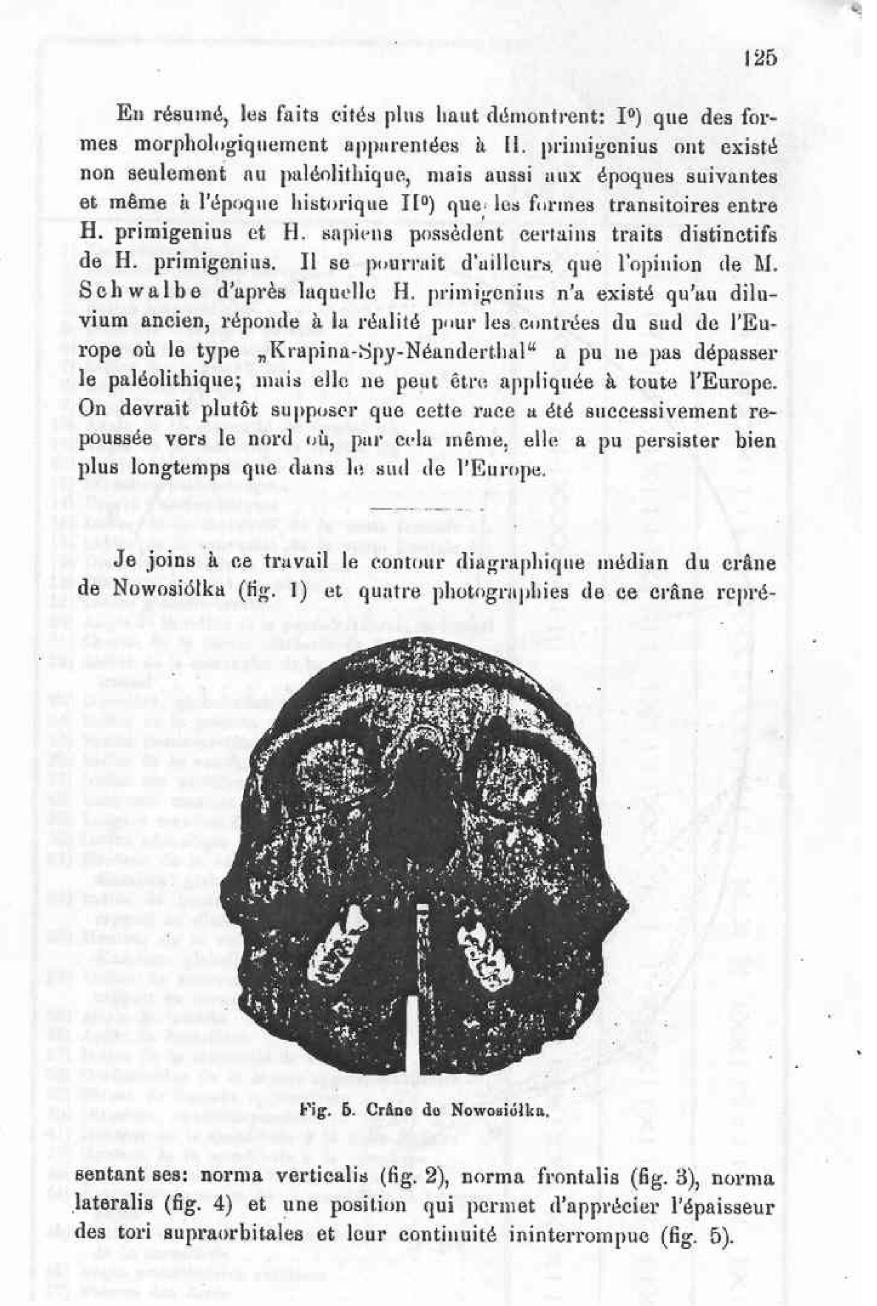
I attach to this work the medien diagraphic countour of the Nowosiolka skull (fig. 1) and four pictures of this skull representing it’s norma verticalis (fig 2), norma frontialis ( fig. 3), normal lateralis (fig. 4) and a position which permits the appreciation of the tori supraorbitalis and their uninterrupted continuum (fig 5).
I wish to express my sincere thanks to M. Schwalbe, L Kollmann, A. Rauber, and K. Maska who have made available several of their precious works. I thank also M. Z. Weyberg for the excellent reproductions of the Nowosiolka skull.
Anthropology Laboratory and Museum of Industry and Agriculture of Varsovie.
It has been brought to my attention that Mike Arehart in Idaho was the individual who found this original manuscript that so many have looked for for so long. Mike, I know I say “Thanks!” for a LOT of people – I’ve been very surprised by the interest this article has generated.
Because of copyright restrictions, only portions of the original manuscript are provided here until I pin down how much of the original I can present here. So, I chose the most pertinent pages.














 Your instructor, Ian Juby, has taught science and technology for 32 years. A robotics engineer by trade with an ambition for life-long learning, he has taught thousands of people from ages 8 to 65 at a premiere summer science camp as well as the high school and collegiate level.
Your instructor, Ian Juby, has taught science and technology for 32 years. A robotics engineer by trade with an ambition for life-long learning, he has taught thousands of people from ages 8 to 65 at a premiere summer science camp as well as the high school and collegiate level.